1. NF-κB signaling pathway
NF-κB 通路作用機制
當處于激活狀態時,NF-κB 位于細胞質中且與抑制蛋白 IκBα 形成復合體。通過內在膜受體的介導,一些胞外信號物質可激活一種稱為 IκB 激酶(IKK)的酶。IKK 轉而磷酸化 IκBα 蛋白,這將導致后者的泛素化,使得 IκBα 從 NF-κB 上脫離下來,最終 IκBα 被蛋白酶體所降解。被激活的 NF-κB 接下來轉移到細胞核內,在這里會結合到 DNA 上被稱為反應元件(RE)的特異性序列上。
DNA/NF-κB 復合體接下來會招募其它蛋白,如輔激活物與 RNA 聚合酶,這些蛋白將下游的 DNA 轉錄為 mRNA 并轉而被翻譯為蛋白質,這些蛋白最終導致細胞功能發生改變。
NF-κB 的調控失常與癌癥、炎癥和自身免疫病、感染性休克、病毒感染以及免疫發育異常有關。


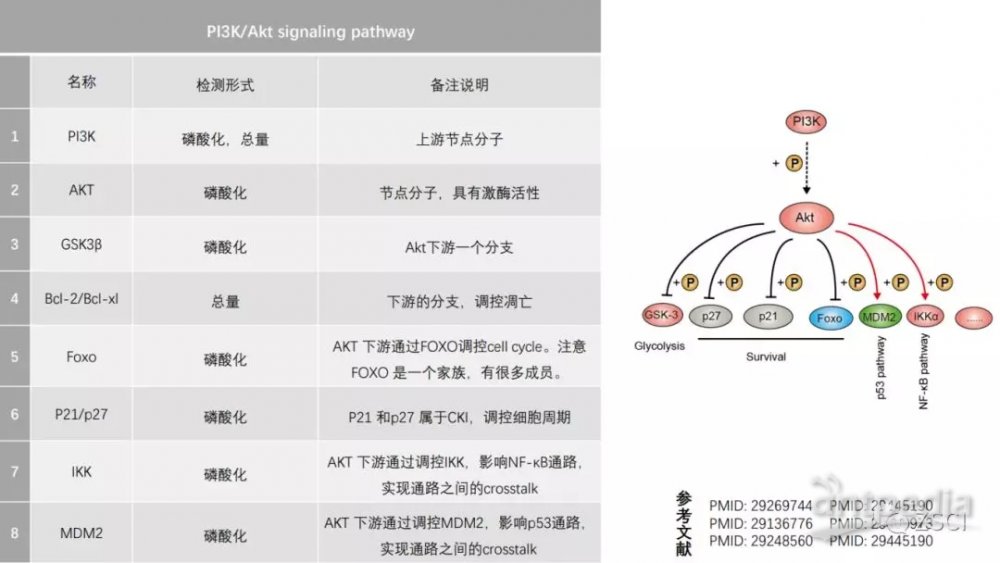
2. PI3K/Akt signaling pathway
PI3K/AKT 通路作用機制:
PI3K 的活化: PI3K 有很多種類別,不過只有第 I 類能夠響應生長刺激而磷酸化脂質。I 類的 PI3K 是異源二聚體,其亞基為 p85(調控)與 p110(催化)。
磷脂酰肌醇的形成:活化的 PI3K 催化磷酸基團到磷酸肌醇的肌醇環上的 3『-OH 位置的加成反應,反應有三種脂質產物, PI (3) P, PI (3,4) P2 和 PI (3,4,5) P3,這些磷酸化的脂質被錨定在細胞膜上,并且可以直接結合細胞內包含 PH 或 FYVE 結構域的蛋白。
Akt 的活化:Akt 以非活化構象駐留在細胞質中,直到細胞被激活,Akt 易位到細胞膜上 Akt 的 PH 作用域對于第二信使 PI (3,4,5) P3 相比其他的磷脂酰肌醇具有更高的親和力。

3. MAPK signaling pathway
MAPK 通路作用機制:
胞外信號→膜受體→RAS→MAP3K→MAP2K→MAPK 然后再進一步活化其他下游靶基因。
MAPK 主要由四個亞家族,分別是:
細胞外信號調節激酶 (extracellular-signalregulated protein kinase, ERK)
p38 絲裂原活化蛋白激酶 (p38 MAPK)
c-Jun 氨基末端激酶 (JNK)
細胞外信號調節激酶 5 (ERK5)
這幾種 MAPK 亞家族參與的信號轉導通路司職不同的功能,如 ERK 調控細胞生長和分化,JNK 和 p38 MAPK 信號通路在炎癥和細胞凋亡等應激反應中發揮重要作用。


4. JAK/STAT signaling pathway
JAK/STAT 通路由三個主要組成部分:
接受信號的酪氨酸激酶相關受體
傳遞信號的酪氨酸激酶 JAK
產生效應通路的轉錄因子 STAT
JAK/STAT 通路調控機制:
來自干擾素、白細胞介素、生長因子或其它化學信使的信號可以激活此受體;
這激活了 JAK 的激酶功能,導致對其自身的磷酸化(磷酸基團作為蛋白質上的開關);
接下來 STAT 蛋白結合到被磷酸化的受體上,在此 STAT 被 JAK 磷酸化;
被磷酸化的 STAT 蛋白結合到另一個被磷酸化的 STAT 蛋白上(二聚化)并易位到細胞核中;
在細胞核中,它結合到 DNA 上并啟動轉錄那些響應 STAT 的基因。


5. TGFβ/SMAD signaling pathway
TGFβ/SMAD 通路調控機制:
TGF-β 雙聚體會結合到 type II 受體
type II 受體會吸引并磷酸化 type I 受體
磷酸化后的 type I 受體吸引并磷酸化 regulated SMAD(R-SMAD)
磷酸化后的 R-SMAD 會結合上 common SMAD(coSMAD、SMAD4)并形成異元二聚體(heterodimeric complex)
該異元二聚體會進入細胞核中作為多種基因表現的轉譯因子,包括利用 8 種途徑活化促分裂蛋白質激酶(mitogen-activated protein kinase)的產生,進而引發細胞凋亡。
而 SMAD 途徑本身被回饋作用所調控,SMAD6 與 SMAD7 可結合上 type I 受體,造成該受體無法與 R-SMAD 結合導致訊息中斷。


6. Wnt/β-catenin signaling pathway
Wnt/β-catenin 通路調控機制:
當 Wnt 蛋白與細胞表面 Frizzled 受體家族結合后的一系列反應,包括 Dishevelled 受體家族蛋白質的激活及最終細胞核內 β-catenin 水平的變化。
Dishevelled (DSH) 是細胞膜相關 Wnt 受體復合物的關鍵成分,它與 Wnt 結合后被激活,并抑制下游蛋白質復合物,包括 axin、GSK- 3、與 APC 蛋白。axin/GSK- 3 /APC 復合體可促進細胞內信號分子 β-catenin 的降解。
當「β-catenin 降解復合物」被抑制后,胞漿內的 β-catenin 得以穩定存在,部分 β-catenin 進入細胞核與 TCF/LEF 轉錄因子家族作用并促進特定基因的表達。


7. Notch signaling pathway
Notch 通路調控機制:
Notch 蛋白橫穿過細胞膜,部分在細胞內而部分在細胞外。配體蛋白結合到胞外域后誘導蛋白切斷并釋放胞內域,胞內域進而進入細胞膜并調控基因表達。
Notch 信號通路提升了神經增殖過程中的增殖信號,而其活性被 Numb 所抑制,以促進神經分化。他在胚胎發育中起到重要的調控作用。


8. Hedgehog signaling pathway
Hedgehog 通路調控機制:
Hh 信號傳遞受靶細胞膜上兩種受體 Patched (Ptc) 和 Smoothened (Smo) 的控制。受體 Ptc 由腫瘤抑制基因 Patched 編碼,是由 12 個跨膜區的單一肽鏈構成,能與配體直接結合,對 Hh 信號起負調控作用。
受體 Smo 由原癌基因 Smothened 編碼,與 G 蛋白偶聯受體同源,由 7 個跨膜區的單一肽鏈構成,N 端位于細胞外,C 端位于細胞內,跨膜區氨基酸序列高度保守,C 末端的絲氨酸與蘇氨酸殘基為磷酸化部位,蛋白激酶催化時結合磷酸基團。該蛋白家族成員只有當維持全長時才有轉錄啟動子的功能,啟動下游靶基因的轉錄。

